Universality of Inheritance and Transmission by Inheritance.—Special Evidences of the same.—Human Beings with four, six, or seven Fingers and Toes.—Porcupine Men.—Transmission of Diseases, especially Diseases of the Mind.—Original Sin.—Hereditary Monarchies.—Hereditary Aristocracy.—Hereditary Talents and Mental Qualities.—Material Causes of Transmission by Inheritance.—Connection between Transmission by Inheritance and Propagation.—Spontaneous Generation and Propagation.—Non-sexual or Monogonous Propagation.—Propagation by Self-Division.—Monera and Amœbæ.—Propagation by the formation of Buds, by the formation of Germ-Buds, by the formation of Germ-Cells.—Sexual or Amphigonous Propagation.—Formation of Hermaphrodites.—Distinction of Sexes, or Gonochorism.—Virginal Breeding, or Parthenogenesis.—Material Transmission of Peculiarities of both Parents to the Child by Sexual Propagation.—Difference between Transmission by Inheritance in Sexual and in Asexual Propagation.
The reader has, in the last chapter, become acquainted with natural selection according to Darwin’s theory, as the constructive force of nature which produces the different forms of animal and vegetable species. By natural selection we understand the interaction which takes place in the struggle for life between the transmission by inheritance and the mutability of organisms, between two physiological functions which are innate in all animals and plants,176 and which may be traced to other processes of life—the functions of propagation and nutrition. All the different forms of organisms, which people are usually inclined to look upon as the products of a creative power, acting for a definite purpose, we, according to the Theory of Selection, can conceive as the necessary productions of natural selection, working without a purpose,—as the unconscious interaction between the two properties of Mutability and Hereditivity. Considering the importance which accordingly belongs to these vital properties of organisms, we must examine them a little more closely, and employ a chapter with the consideration of Transmission by Inheritance. (Gen. Morph. ii. 170-191.)
Strictly speaking, we must distinguish between Hereditivity (Transmissivity) and Inheritance (Transmission). Hereditivity is the power of transmission, the capability of organisms to transfer their peculiarities to their descendants by propagation. Transmission by Inheritance, or Inheritance simply, on the other hand, denotes the exercise of the capability, the actual transmission.
Hereditivity and Transmission by Inheritance are such universal, everyday phenomena, that most people do not heed them, and but few are inclined to reflect upon the operation and import of these phenomena of life. It is generally thought quite natural and self-evident that every organism should produce its like, and that children should more or less resemble their parents. Heredity is usually only taken notice of and discussed in cases relating to some special peculiarity, which appears for the first time in a human individual without having been inherited, and then is transmitted to his descendants. It shows177 itself in a specially striking manner in the case of certain diseases, and in unusual and irregular (monstrous) deviations from the usual formation of the body.
Among these cases of the inheritance of monstrous deviations, those are specially interesting which consist in an abnormal increase or decrease of the number five in the fingers or toes of man. It is not unfrequently observed in families through several generations, that individuals have six fingers on each hand, or six toes on each foot. Less frequent is the number of four or seven fingers or toes. The unusual formation arises at first from a single individual who, from unknown causes, is born with an excess of the usual number of fingers and toes, and transmits these, by inheritance, to a portion of his descendants. In one and the same family it has happened that, throughout three, four, or more generations, individuals have possessed six fingers and toes. In a Spanish family there were no less than forty individuals distinguished by this excess. The transmission of the sixth finger or toe is not permanent or enduring in all cases, because six-fingered people always intermarry again with those possessing five fingers. If a six-fingered family were to propagate by pure in-breeding, if six-fingered men were always to marry six-fingered women, this characteristic would become permanent, and a special six-fingered human race would arise. But as six-fingered men usually marry five-fingered women, and vice versâ, their descendants for the most part show a very mixed numerical relation, and finally, after the course of some generations, revert again to the normal number of five. Thus, for example, among eight children of a six-fingered father and a five-fingered mother, two children may have on both hands and feet six fingers178 and toes, four children may have a mixed number, and two children may have the usual number of five on both hands and feet. In a Spanish family, each child except the youngest had the number six on both hands and feet; the youngest, only, had the usual number on both hands and feet, and the six-fingered father of the child refused to recognize the last one as his own.
The power of inheritance, moreover, shows itself very strikingly in the formation and colour of the human skin and hair. It is well known how exactly the nature of the complexion in many families—for instance, a peculiar soft or rough skin, a peculiar luxuriance of the hair, a peculiar colour and largeness of the eyes—is transmitted through many generations. In like manner, peculiar local growths or spots on the skin, the so-called moles, freckles, and other accumulations of pigment which appear in certain places, are frequently transmitted through several generations so exactly, that in the descendants they appear on the same spots on which they existed in the parents. The porcupine men of the Lambert family, who lived in London last century, are especially celebrated. Edward Lambert, born in 1717, was remarkable for a most unusual and monstrous formation of the skin. His whole body was covered with a horny substance, about an inch thick, which rose in the form of numerous thorn-shaped and scale-like processes, more than an inch long. This monstrous formation of the outer skin, or epidermis, was transmitted by Lambert to his sons and grandsons, but not to his granddaughters. The transmission in this instance remained in the male line, as is often the case. In like manner, an excessive development of fat in certain parts of the body is often transmitted179 only in the female line. I scarcely need call to mind how exactly the characteristic formation of the face is transmitted by inheritance; sometimes it remains within the male, sometimes within the female line; sometimes it is blended in both.
The phenomena of transmission by inheritance of pathological conditions, especially of the different forms of human diseases, are very instructive and generally known. Diseases of the respiratory organs, the glands, and of the nervous system, are specially liable to be transmitted by inheritance. Very frequently there suddenly appears in an otherwise healthy family a disease until then unknown among them; it is produced by external causes, by conditions of life causing disease. This disease, brought about in an individual by external cause, is propagated and transmitted to his descendants, and some or all of them then suffer from the same disease. In case of diseases of the lungs, for instance in consumption, this sad transmission by inheritance is well known, and it is the same with diseases of the liver, with syphilis, and diseases of the mind. The latter are specially interesting. Just as peculiar characteristic features of man—pride, ambition, frivolity, etc.—are transmitted to the descendants strictly by inheritance, so too are the peculiar abnormal manifestations of mental activity, which are usually called fixed ideas, despondency, imbecility, and generally “diseases of the mind.” This distinctly and irrefragably shows that the soul of man, just as the soul of animals, is a purely mechanical activity, the sum of the molecular phenomena of motion in the particles of the brain, and that it is transmitted by inheritance, together with its substratum, just as every other quality of the body is materially transmitted by propagation.180
When this exceedingly important and undeniable fact is mentioned, it generally causes great offence, and yet in reality it is silently and universally acknowledged. For upon what else do the ideas of “hereditary sin,” “hereditary wisdom,” and “hereditary aristocracy,” etc., rest than upon the conviction that the quality of the human mind is transmitted by propagation—that is, by a purely material process—through the body, from the parents to the descendants? The recognition of this great importance of transmission by inheritance is shown in a number of human institutions, as for example, among many nations in the division into castes, such as the castes of warriors, castes of priests, and castes of labourers, etc. It is evident that the institution of such castes originally arose from the notion of the great importance of hereditary distinctions possessed by certain families, which it was presumed would always be transmitted by the parents to the children. The institution of an hereditary aristocracy and an hereditary monarchy is to be traced to the notion of such a transmission of special excellencies. However, it is unfortunately not only virtues, but also vices that are transmitted and accumulated by inheritance; and if, in the history of the world, we compare the different individuals of the different dynasties, we shall everywhere find a great number of proofs of the transmission of qualities by inheritance, but fewer of transmissions of virtues than of vices. Look only, for example, at the Roman emperors, at the Julii and the Claudii, or at the Bourbons in France, Spain, and Italy!
In fact, scarcely anywhere could we find such a number of striking examples of the remarkable transmission of bodily and mental features by inheritance, as in the history181 of the reigning houses in hereditary monarchies. This is specially true in regard to the diseases of the mind previously mentioned. It is in reigning families that mental disorders are hereditary in an unusual degree. Thus Esquirol, distinguished for his knowledge of mental diseases, proved that the number of insane individuals in the reigning houses was, in proportion to the number among the ordinary population, as 60 to 1; that is, that disorders of the brain occur 60 times more frequently in the privileged families of the ruling houses than among ordinary people. If equally accurate statistics were made of the hereditary nobility, the result would probably be that here also we should find an incomparably larger contingent of mental diseases than among the common, ignoble portion of mankind. This phenomenon can scarcely astonish us if we consider what injury these privileged castes inflict upon themselves by their unnatural, one-sided education, and by their artificial separation from the rest of mankind. By this means many dark sides of human nature are specially developed and, as it were, artificially bred, and, according to the laws of transmission by inheritance, are propagated through series of generations with ever-increasing force and dominance.
It is sufficiently obvious from the history of nations how in successive generations of many dynasties, for example, of the princes of Saxon Thuringia and of the Medici, the noble solicitude for the most perfect human accomplishments in science and art were retained and transmitted from father to son; and how, on the other hand, in many other dynasties, for centuries a special partiality for the profession of war, for the oppression of human freedom, and for other rude acts of violence, have been hereditary. In like182 manner talents for special mental activities are transmitted in many families for generations, as, for instance, talent for mathematics, poetry, music, sculpture, the investigation of nature, philosophy, etc. In the family of Bach there have been no less than twenty-two eminent musicians. Of course the transmission of such peculiarities of mind depends upon the material process of reproduction, as does the transmission of mental qualities in general. In this case again, the vital phenomenon, the manifestation of force (as everywhere in nature), is directly connected with definite relations in the admixture of the material components of the organism. It is this definite proportion and molecular motion of matter which is transmitted by generation.
Now, before we examine the numerous, and in some cases most interesting and important, laws of transmission by inheritance, let us make ourselves acquainted with the actual nature of the process. The phenomena of transmission by inheritance are generally looked upon as something quite mysterious, as peculiar processes which cannot be fathomed by natural science, and the causes and actual nature of which cannot be understood. It is precisely in such a case that people very generally assume supernatural influences. But even in the present state of our physiology it can be proved with complete certainty that all the phenomena of inheritance are entirely natural processes, that they are produced by mechanical causes, and that they depend on the material phenomena of motion in the bodies of organisms, which we may consider as a part of the phenomena of propagation. All the phenomena of Heredity and the laws of Transmission by Inheritance can be traced to the material process of Propagation.
183Every organism, every living individual, owes its existence either to an act of unparental or Spontaneous Generation (Generatio Spontanea, Archigonia), or to an act of Parental Generation or Propagation (Generatio Parentalis, Tocogonia). In a future chapter we shall have to consider Spontaneous Generation, or Archigony. At present we must occupy ourselves with Propagation, or Tocogony, a closer examination of which is of the utmost importance for understanding transmission by inheritance. Most of my readers probably only know those phenomena of Propagation which are seen universally in the higher plants and animals, the processes of Sexual Propagation, or Amphigony. The processes of Non-sexual Propagation, or Monogony, are much less generally known. The latter, however, are far more suited to throw light upon the nature of transmission by inheritance in connection with propagation.
For this reason, we shall first consider only the phenomena of non-sexual or monogonic propagation (Monogonia). This appears in a variety of different forms, as for example, self-division, formation of buds, the formation of germ-cells or spores (Gen. Morph. ii. 36-58). It will be most instructive, first, to examine the propagation of the simplest organisms known to us, which we shall have to return to later, when considering the question of spontaneous generation. These very simplest of all organisms yet known, and which, at the same time, are the simplest imaginable organisms, are the Monera living in water; they are very small living corpuscles, which, strictly speaking, do not at all deserve the name of organism. For the designation “organism,” applied to living creatures, rests upon the idea that every living natural body is composed 184 of organs, of various parts, which fit into one another and work together (as do the different parts of an artificial machine), in order to produce the action of the whole. During late years we have become acquainted with Monera, organisms which are, in fact, not composed of any organs at all, but consist entirely of shapeless, simple, homogeneous matter. The entire body of one of these Monera, during life, is nothing more than a shapeless, mobile, little lump of mucus or slime, consisting of an albuminous combination of carbon. Simpler or more imperfect organisms we cannot possibly conceive.
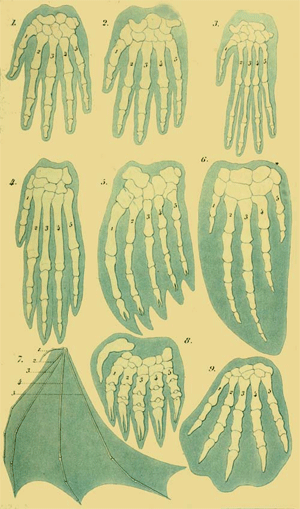
The first complete observations on the natural history of a Moneron (Protogenes primordialis) were made by me at Nice, in 1864. Other very remarkable Monera I examined later (1866) in Lanzarote, one of the Canary Islands, and in 1867 in the Straits of Gibraltar. The complete history of one of these Monera, the orange-red Protomyxa aurantiaca, is represented in Plate I, and its explanation is given in the Appendix. I have found some curious Monera also in the North Sea, off the Norwegian coast, near Bergen. Cienkowski has described (1865) an interesting Moneron from fresh waters, under the name of Vampyrella. But perhaps the most remarkable of all Monera was discovered by Huxley, the celebrated English zoologist, and called Bathybius Hæckelii. “Bathybius” means, living in the deep. This wonderful organism lives in immense depths of the ocean, which are over 12,000—indeed, in some parts 24,000 feet below the surface, and which have become known to us within the last ten years, through the laborious investigations made by the English. There, among the numerous Polythalamia and 185 Radiolaria which inhabit the fine calcareous mud of these abysses, the Bathybius is found in great quantities, sometimes in the shape of roundish, formless lumps of mucus, sometimes in the form of a network of mucus, covering fragments of stone and other objects. Small particles of chalk are frequently embedded in these mucous gelatinous masses, and are, perhaps, products of their secretion. The entire body of this remarkable Bathybius consists solely of shapeless plasma, or protoplasm, as in the case of the other Monera—that is, it consists of the same albuminous combination of carbon, which in infinite modifications is found in all organisms, as the essential and never-failing seat of the phenomena of life. I have given a detailed description and drawing of the Bathybius and other Monera in my “Monographie der Moneren,” 1870,(15) from which the drawing in Fig. 9 is taken.
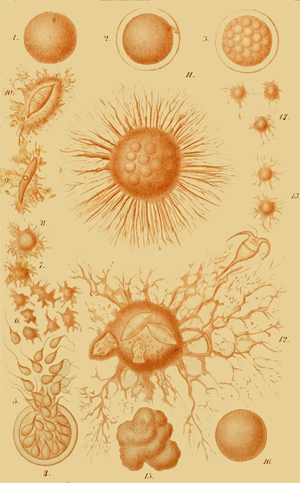
Life history of a simplest organism. Pl. I.
Life history of a simplest organism.
E. Haeckel del. Prototmyxa aurantiaca. Lagesse sc.
In a state of rest most Monera appear as small globules of mucus or slime, invisible, or nearly so, to the naked eye; they are at most as large as a pin’s head. When the Moneron moves itself, there are formed on the upper surface of the little mucous globule, shapeless, fingerlike processes, or very fine radiated threads; these are the so-called false feet, or pseudopodia. The false feet are simple, direct continuations of the shapeless albuminous mass, of which the whole body consists. We are unable to perceive different parts in it, and we can give a direct proof of the absolute simplicity of the semi-fluid mass of albumen, for with the aid of the microscope we can follow the Moneron as it takes in nourishment. When small particles suited for its nourishment—for instance, small particles of decayed organic bodies or microscopic plants and infusoria—accidentally 186 come into contact with the Moneron, they remain hanging to the sticky semi-fluid globule of mucus, and here create an irritation, which is followed by a strong afflux of the mucous substance, and, in consequence, they become finally completely inclosed by it, or are drawn into the body of the Moneron by displacement of the several albuminous particles, and are there digested, being absorbed by simple diffusion (endosmosis).
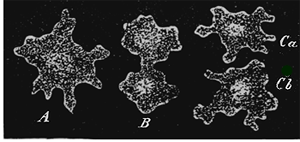
Propagation of the simplest organism, a Moneron, by self-division.
Fig. 1.—Propagation of the simplest organism, a Moneron, by self-division. A. The entire Moneron, a Protamœba. B. It falls into two halves by a contraction in the middle. C. Each of the two halves has separated from the other, and now represents an independent individual.
Just as simple as the process of nutrition is the propagation of these primitive creatures, which in reality we can neither call animals nor plants. All Monera propagate themselves only in an asexual manner by monogony; and in the simplest case, by that kind of monogony which we place at the head of the different forms of propagation, that is, by self-division. When such a little globule, for example a Protamœba or a Protogenes, has attained a certain size by the assimilation of foreign albuminous matter, it falls into two pieces; a pinching in takes place, contracting the middle of the globule on all sides, and finally leads to the separation of the two halves (compare Fig. 1). Each half187 then becomes rounded off, and now appears as an independent individual, which commences anew the simple course of the vital phenomena of nutrition and propagation. In other Monera (Vampyrella), the body in the process of propagation does not fall into two, but into four equal pieces, and in others, again (Protomonas, Protomyxa, Myxastrum), at once into a number of small globules of mucus, each of which again, by simple growth, becomes like the parent body. Here it is evident that the process of propagation is nothing but a growth of the organism beyond its own individual limit of size.
The simple method of propagation of the Moneron by self-division is, in reality, the most universal and most widely spread of all the different modes of propagation; for by the same simple process of division, cells also propagate themselves. Cells are those simple organic individuals, a large number of which constitute the bodies of most organisms, the human body not excepted. With the exception of the organisms of the lowest order, which have not even the perfect form of a cell (Monera), or during life only represent a single cell (many Protista and single-celled plants), the body of every organic individual is composed of a great number of cells. Every organic cell is to a certain degree an independent organism, a so-called “elementary organism,” or an “individual of the first order.” Every higher organism is, in a measure, a society or a state of such variously shaped elementary individuals, variously developed by division of labour.(39) Originally every organic cell is only a single globule of mucus, like a Moneron, but differing from it in the fact that the homogeneous albuminous substance has separated itself into two different parts, a firmer albuminous 188 body, the cell-kernel (nucleus), and an external, softer albuminous body, the cell-substance or body (protoplasma). Besides this, many cells later on form a third (frequently absent) distinct part, inasmuch as they cover themselves with a capsule, by exuding an outer pellicle or cell-membrane (membrana). All other forms of cells, besides these, are of subordinate importance, and are of no further interest to us here.
Every organism composed of many cells was originally a single cell, and it becomes many-celled owing to the fact that the original cell propagates itself by self-division, and that the new individual cells originating in this manner remain together, and by division of labour form a community or a state. The forms and vital phenomena of all many-celled organisms are merely the effect or the expression of all the forms and vital phenomena of all the individual cells of which they are composed. The egg, from which most animals and plants are developed, is a simple cell.
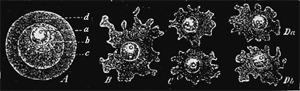
Propagation of a single-celled organism
Fig. 2.—Propagation of a single-celled organism, Amœba sphærococcus, by self-division. A. The enclosed Amœba, a simple globular cell consisting of a lump of protoplasm (c), which contains a kernel (b) and a kernel speck (a), and is surrounded by a cell-membrane or capsule. B. The free Amœba, which has burst and left the cyst or cell-membrane. C. It begins to divide by its kernel forming two kernels, and by the cell-substance between the two becoming contracted. D. The division is completed by the cell-substance likewise falling into two halves (Da and Db).
The single-celled organisms, that is, those which during life retain the form of a single cell, for example the Amœbæ, as a rule propagate themselves in the simplest way by self-division. This process differs from the previously described self-division of the Moneron only in the fact that at the commencement the firmer cell-kernel (nucleus) falls into two halves, by a pinching in at its middle. The two young kernels separate from each other and act now as two distinct centres of attraction upon the surrounding softer albuminous matter, that is, the cell-substance (protoplasma). By this process finally the latter also divides into two halves, and there now exist two new cells, which are like the mother cell. If the cell was surrounded by a membrane, this either does not divide at all, as in the case of egg-cleavage (Fig. 3, 4), or it passively follows the active pinching in of the protoplasm; or, lastly, every new cell exudes a new membrane for itself.
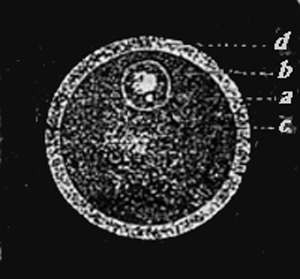
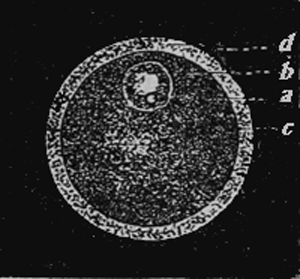
Egg of a mammal (a simple cell).
Fig. 3.—Egg of a mammal (a simple cell). a. The small kernel speck or nucleolus (the so-called germ-spot of the egg). b. Kernel or nucleus (the so-called germ-bladder of the egg). c. Cell-substance or protoplasm (the so-called yolk of the egg). d. Cell-capsule or membrane (membrane of the yolk) of the egg; called in mammals, on account of its transparency, Membrana pellucida.
The non-independent cells which remain united in communities or states, and thus constitute the body of higher organisms, are propagated in the same manner as are independent single-celled organisms, for example, Amœba (Fig. 2). Just as in that case, the cell with which most animals and plants commence their individual existence, namely, the egg, multiplies itself by simple division. When an animal, for instance a mammal (Fig. 3, 4), develops out of an190 egg, this process of development always begins by the simple egg-cell (Fig. 3) forming an accumulation of cells (Fig. 4) by continued self-division. The outer covering, or cell membrane, of the globular egg remains undivided. First, the cell-kernel of the egg (the so-called germinal vesicle) divides itself into two kernels, then follows the cell-substance (the yolk of the egg) (Fig. 4 A). In like manner, the two cells, by continued self-division, separate into four (Fig. 4 B), these into eight (Fig. 4 C), into sixteen, thirty-two, etc., and finally there is produced a globular mass of very numerous little cells (Fig. 4 D). These now, by further increase and heterogeneous development (division of labour), gradually build up the compound many-celled organism. Every one of us, at the commencement of our individual development, has undergone the very same process as that represented in Fig. 4. The egg of a mammal—represented in Fig. 3, and its development in Fig. 4—might as well be that of a man, as of an ape, dog, horse, or any other placental mammal.

First commencement of the development of a mammal’s egg.
Fig. 4.—First commencement of the development of a mammal’s egg, the so-called “cleavage of the egg” (propagation of the egg-cell by repeated self-division). A. The egg, by the formation of the first furrow, falls into two cells. B. These separate by division into four cells. C. The latter have divided into eight cells. D. By repeated division a globular accumulation of numerous cells has arisen.
Now, when one examines this simplest form of propagation, this self-division, it surely cannot be considered wonderful that the products of the division of the original organism should possess the same qualities as the parental individual. For they are parts or halves of the parental organism, and the matter or substance in both halves is the same, and as both the young individuals have received an equal amount and the same quality of matter from the parent individual, one can but consider it natural that the vital phenomena, the physiological qualities should be the same in both children. In fact, in regard to their form and substance, as well as to their vital phenomena, the two produced cells can in no respect be distinguished from one another, or from the mother cell. They have inherited from her the same nature.
But this same simple propagation by self-division is not only confined to simple cells—it is the same also in the higher many-celled organisms; for example, in the coral zoophytes. Many of them which exhibit a high complexity of composition and organization, nevertheless, propagate themselves by simple division. In this case the whole organism, with all its organs, falls into two equal halves as soon as by growth it has attained a certain size. Each half again develops itself, by growth, into a complete individual. Here, again, it is surely self-evident that the two products of division will share the qualities of the parental organism, as they themselves are in fact halves of that parent.
Next to propagation by division we come to propagation by the formation of buds. This kind of monogony is exceedingly widely spread. It occurs both in the case of simple cells (though not frequently) and in the higher organisms 192 composed of many cells. The formation of buds is universal in the vegetable kingdom, less frequent in the animal kingdom. However, here also it occurs in the tribe of Plant-like Animals, especially among the Coral Zoophytes, and among the greater portion of the Hydroid Polyps very frequently, further also among some worms (Planarian Worms, Ring-Worms, Moss Animals, Tunicates). Most branching animal-trees or colonies, which are exceedingly like branching plants, arise like those plants, by the formation of buds.
Propagation by the formation of buds (Gemmatio) is essentially distinguished from propagation by division, in the fact that the two organisms thus produced by budding are not of equal age, and therefore at first are not of equal value, as they are in the case of division. In division we cannot clearly distinguish either of the two newly produced individuals as the parental, that is as the producer, because, in fact, both have an equal share in the composition of the original parental individual. If, on the other hand, an organism sends out a bud, then the latter is the child of the former. The two individuals are of unequal size and of unequal form. If, for instance, a cell propagates itself by the formation of buds, we do not see the cell fall into two equal halves, but there appears at one point of it a protuberance, which becomes larger and larger, more or less separates itself from the parental cell, and then grows independently. In like manner we observe in the budding of a plant or animal, that a small local growth arises on a part of the mature individual, which growth becomes larger and larger, and likewise more or less separates itself from the parental organism by an independence in its growth. The bud, after193 it has attained a certain size, may either completely separate itself from the parental individual, or it may remain connected with it and form a stock or colony, whilst at the same time its life may be quite independent of that of its parent. While the growth which starts the propagation, in the case of self-division, is a total one affecting the whole body, it is in the formation of buds only partial, affecting merely a portion of the parental organism. But here, also, the bud—the newly-produced individual which remains so long most directly connected with the parental organism, and which proceeds from it—retains the essential qualities and the original tendency of development of its parent.
A third mode of non-sexual propagation, that of the formation of germ-buds (Polysporogonia), is intimately connected with the formation of buds. In the case of the lower, imperfect organisms, among animals, especially in the case of the Plant-like animals and Worms, we very frequently find that in the interior of an individual composed of many cells, a small group of cells separates itself from those surrounding it, and that this small isolated group gradually develops itself into an individual, which, becomes like the parent, and sooner or later comes out of it. Thus, for example, in the body of the Fluke-worms (Trematodes) there often arise numerous little bodies consisting of many cells, that is germ-buds, or polyspores, which, at an early stage separate themselves completely from the parent body, and leave it when they have attained a certain stage of development.
The formation of germ-buds is evidently but little different from real budding. But, on the other hand, it is connected with a fourth kind of non-sexual propagation, which almost194 forms a transition to sexual reproduction, namely, the formation of germ-cells (Monosporogonia), which is often briefly called formation of spores (sporogonia). In this case it is no longer a group of cells, but a single cell, which separates itself from the surrounding cells in the interior of the producing organism, and which only becomes further developed after it has come out of its parent. After this germ-cell, or monospore (or, briefly, spore), has left the parental individual, it multiplies by division, and thus forms a many-celled organism, which by growth and gradual development attains the hereditary qualities of the parental organism. This occurs very generally among lower plants (Cryptogama).
Although the formation of germ-cells very much resembles the formation of germ buds, it evidently and very essentially differs from the latter, and also from the other forms of non-sexual propagation which have previously been mentioned, by the fact that only a very small portion of the producing organism takes part in the propagation and, accordingly, in the transmission by inheritance. In the case of self-division, where the whole organism falls into two halves, in the formation of buds, where a considerable portion of the whole body, already more or less developed, separates from the producing individual, we easily understand that the forms and vital phenomena should be the same in the producing and produced organism. It is much more difficult to understand in the formation of germ-buds, and more difficult still in the formation of germ-cells, how this very small, quite undeveloped portion of the body, this group of cells, or this single cell, not only directly takes with it certain parental qualities into its independent existence, but also after its195 separation from the parental individual develops into a many-celled body, and in this repeats the forms and vital phenomena of the original producing organism. This last form of monogonic propagation—that of the germ cells, or spore-formation—leads us directly to a form of propagation which is the most difficult of all to explain, namely, sexual propagation.
Sexual or amphigonic propagation (Amphigonia) is the usual method of propagation among all higher animals and plants. It is evident that it has only developed, at a very late period of the earth’s history, from non-sexual propagation, and apparently in the first instance from the method of propagation by germ-cells. In the earliest periods of the organic history of the earth, all organisms propagated themselves in a non-sexual manner, as numerous lower organisms still do, especially all those which are at the lowest stage of organization, and which, strictly speaking, can be considered neither as animals nor as plants, and which therefore, as primary creatures, or Protista, are best excluded from both the animal and vegetable kingdoms. In the case of the higher animals and plants, the increase of individuals, as a rule, is at present brought about in the majority of cases by sexual propagation.
In all the chief forms of non-sexual propagation mentioned above—in fission, in the formation of buds, germ buds, and germ cells—the separated cell or group of cells was able by itself to develop into a new individual, but in the case of sexual propagation the cell must first be fructified by another generative substance. The fructifying male sperm must first mix with the female germ-cell (the egg) before the latter can develop into a new individual. These two196 different generative substances, the male sperm and the female egg, are either produced by one and the same individual hermaphrodite (Hermaphroditismus), or by two different individuals (sexual separation, Gonochorismus) (Gen. Morph. ii. 58, 59).
The simpler and more ancient form of sexual propagation is through double-sexed individuals (Hermaphroditismus). It occurs in the great majority of plants, but only in a minority of animals, for example, in the garden snails, leeches, earth-worms, and many other worms. Every single individual among hermaphrodites produces within itself materials of both sexes—eggs and sperm. In most of the higher plants every blossom contains both the male organ (stamens and anther) and the female organs (style and germ). Every garden snail produces in one part of its sexual gland eggs, and in another part sperm. Many hermaphrodites can fructify themselves; in others, however, copulation and reciprocal fructification of both hermaphrodites is necessary for causing the development of the eggs. This latter case is evidently a transition to sexual separation.
Sexual separation (Gonochorismus,) which characterizes the more complicated of the two kinds of sexual reproduction, has evidently been developed from the condition of hermaphroditism at a late period of the organic history of the world. It is at present the universal method of propagation of the higher animals, and occurs, on the other hand, only in the minority of plants (for example, in many aquatic plants, e.g. Hydrocharis, Vallisneria; and in trees, e.g. Willows, Poplars). Every organic individual, as a non-hermaphrodite (Gonochoristus), produces within itself only197 one of two generative substances, either the male or the female. The female individuals, both in animals and plants, produce eggs or egg-cells. The eggs of plants in the case of flowering plants (Phanerogama), are commonly called “embryo sacs”; in the case of flowerless plants (Cryptogama), “fruit spores.” In animals, the male individual secretes the fructifying sperm (sperma); in plants, the corpuscles, which correspond to the sperm. In the Phanerogama, these are the pollen grains, or flower-dust; in the Cryptogama, a sperm, which, like that of most animals, consists of floating vibratile cells actively moving in a fluid—the zoosperms, spermatozoa, or sperm-cells.
The so-called virginal reproduction (Parthenogenesis) offers an interesting form of transition from sexual reproduction to the non-sexual formation of germ-cells (which most resembles it); it has been demonstrated to occur in many cases among Insects, especially by Siebold’s excellent investigations. In this case germ-cells, which otherwise appear and are formed exactly like egg-cells, become capable of developing themselves into new individuals without requiring the fructifying seed. The most remarkable and most instructive of the different partheno-genetic phenomena are furnished by those cases in which the same germ-cells, according as they are fructified or not, produce different kinds of individuals. Among our common honey bees, a male individual (a drone) arises out of the eggs of the queen, if the egg has not been fructified; a female (a queen, or working bee), if the egg has been fructified. It is evident from this, that in reality there exists no wide chasm between sexual and non-sexual reproduction, but that both modes of reproduction are directly198 connected. The parthenogenesis of Insects must probably be regarded as a relapse from the sexual mode of propagation (possessed by the original parents of the insects) to the earlier condition of non-sexual propagation. (Gen. Morph. ii. 86.) In any case, however, sexual reproduction, both in plants and animals, which seems such a wonderful process, has only arisen at a later date out of the more ancient process of non-sexual reproduction. In both cases heredity is a necessary part of the phenomenon.
In all the different modes of propagation the essential point of the process is invariably a detachment of a portion of the parental organism possessing the capability of leading an individual, independent existence. We may, therefore, in all cases expect, à priori, that the produced individuals—which are, in fact, as is commonly said, “the flesh and blood” of the parents—will receive the vital characteristics and qualities of form which the parental individuals possess. It is simply a larger or smaller quantity of the parental material, in fact of its albuminous protoplasm, or cell-substance, which passes to the produced individual. But together with the material, its vital properties—that is, the molecular motions of the plasma—are transmitted, which then manifest themselves in its form. Inheritance by sexual breeding loses very much of the mysterious and wonderful character which it at first sight possesses for the uninitiated, if we consider the above-mentioned series of the different modes of propagation, and their connection one with another. It at first appears exceedingly wonderful that in the sexual propagation of man, and of all higher animals, the small egg, the minute cell, often invisible to the naked eye, is able to transfer to the produced organism all the qualities199 of the maternal organism, and, no less mysterious, that at the same time the essential qualities of the paternal organism are transferred to the offspring by means of the male sperm, which fructifies the egg-cell by means of a viscid substance in which minute thread-like cells or zoosperms move about. But as soon as we compare the connected stages of the different kinds of propagation, in which the produced organism separates itself more and more as a distinct growth from the parental individual, and more or less early enters upon its independent career; as soon as we consider, at the same time, that the growth and development of every higher organism only depends upon the increase of the cells composing it—that is, upon their simple propagation by division—it becomes quite evident that all these remarkable processes belong to one series.
The life of every organic individual is nothing but a connected chain of very complicated material phenomena of motion. These motions must be considered as changes in the position and combination of the molecules, that is, of the smallest particles of animated matter (of atoms placed together in the most varied manner). The specific, definite tendency of these orderly, continuous, and inherent motions of life depends, in every organism, upon the chemical mingling of the albuminous generative matter to which it owes its origin. In man, as in the case of the higher animals which propagate themselves in a sexual manner, the individual vital motion commences at the moment in which the egg-cell is fructified by the spermatic filaments of the seed, in which process both generative substances actually mix; and here the tendency of the vital motion is determined by the specific, or more200 accurately, by the individual nature of the sperm as well as of the egg. There can be no doubt as to the purely mechanical material nature of this process. But here we stand full of wonder and astonishment before the infinite and inconceivable delicacy of this albuminous matter. We are amazed at the undeniable fact that the simple egg-cell of the maternal organism, and a single paternal sperm-thread, transfer the molecular individual vital motion of these two individuals to the child so accurately, that afterwards the minutest bodily and mental peculiarities of both parents reappear in it.
Here we stand before a mechanical phenomenon of nature of which Virchow, whose genius founded the “cellular pathology,” says with full justice: “If the naturalist cared to follow the custom of historians and preachers, and to clothe phenomena, which are in their way unique, with the hollow pomp of ponderous and sounding words, this would be the opportunity for him; for we have now approached one of those great mysteries of animal nature, which encircle the region of animal life as opposed to all the rest of the world of phenomena. The question of the formation of cells, the question of the excitation of a continuous and equable motion, and, finally, the questions of the independence of the nervous system and of the soul—these are the great problems on which the human mind can measure its strength.” To comprehend the relation of the male and female to the egg-cell is almost as much as to solve all those mysteries. The origin and development of the egg-cell in the mother’s body, the transmission of the bodily and mental peculiarities of the father to it by his seed, touch upon all the questions which the human201 mind has ever raised about man’s existence. And, we add, these most important questions are solved, by means of the Theory of Descent, in a purely mechanical and purely monistic sense!
There can then be no further doubt that, in the sexual propagation of man and all higher organisms, inheritance, which is a purely mechanical process, is directly dependent upon the material continuity of the producing and produced organism, just as is the case in the simplest non-sexual propagation of the lower organisms. However, I must at once take this opportunity of drawing attention to an important difference which inheritance presents in sexual and non-sexual propagation. It is a fact long since acknowledged, that the individual peculiarities of the producing organism are much more accurately transmitted to the produced organism by non-sexual than by sexual propagation. Gardeners have for a long time made use of this fact in many ways. When, for instance, a single individual of a species of tree with stiff, upright branches accidentally produces down-hanging branches, a gardener, as a rule, cannot transmit this peculiarity by sexual, but only by non-sexual propagation. The twigs cut off such a weeping tree and planted as cuttings or slips, afterwards produce trees having likewise hanging branches, as, for example, the weeping willows and beeches. Seedlings, on the other hand, which have been reared out of the seed of such a weeping tree, generally have the original stiff and upright form of branches possessed by their ancestors. The same may be observed in a very striking manner in the so-called “copper-coloured trees,” that is, varieties of trees which are characterized by a red or reddish brown202 colour of the leaves. Off-shoots from such copper-coloured trees (for example, the copper beech), which have been propagated by cuttings in a non-sexual manner, show the peculiar colour and nature of the leaves which distinguished the parental individual, while others reared from seeds of such a copper-coloured tree return to the green-coloured condition of leaf.
This difference in inheritance will seem very natural when we consider that the material connection between the producing and produced individuals is much closer and lasts much longer in non-sexual than in sexual propagation. The special tendency of the molecular motion of life can therefore fix itself much longer and more thoroughly in the filial organism, and be more strictly transmitted by non-sexual than by sexual propagation. All these phenomena, considered in connection, clearly prove that the transmission of bodily and mental peculiarities is a purely material and mechanical process. By propagation a greater or lesser quantity of albuminous particles, and together with them the individual form of motion inherent in these molecules of protoplasm, are transmitted from the parental organism to the offspring. As this form of motion remains continuous, the more delicate peculiarities inherent in the parental organism must sooner or later reappear in the filial organism.